-
|
-
概览
• 感染Sars-CoV-2的K18-hACE2转基因小鼠的神经炎症
• 败血症小鼠模式
• 产品
• 参考文献
相关产品
• CD11b抗体, HS-384 103, 兔多克隆抗体,小鼠特定
• CD11b抗体, HS-384 117,大鼠单克隆抗体,小鼠特定
小胶质细胞是中枢神经系统(CNS)包括脑、脊髓、视网膜和嗅球的重要免疫效应细胞。小胶质细胞功能在于支持中枢神经系统(CNS)的发育,维持内环境稳定和免疫反应。未激活或‘休眠’的小胶质细胞是网状的,并会形成一个横跨中枢神经系统的密集网络。通过高度运动的长细胞过程,它们会积极地筛查微环境,防止稳态的破坏(Kabba et al., 2018)。在未激活的状态下,小胶质细胞可表达标记物IBA-1、CD68、CD11b、CD40、CD45、CD80、CD86、F4/80、TREM-2b、CXCR3 和CCR9 (Bachiller et al., 2018; Jurga et al., 2020)。然而,也发现表达也有区域性的差异。例如,海马体小胶质细胞可表达更高水平F4/80,而CD11b在大脑皮层的小胶质细胞中时常会比脊髓中更低(Bachiller et al., 2018)。
图1:小胶质细胞活化的代表示意图(改编自 Pinto et al., 2020)。在响应病原体或脑中内环境稳态改变时,小胶质细胞活化会引起形态和分子的改变。活化的细胞会部分地收缩其过程,从网状到中间的‘浓密’,到最终的具有吞噬功能、状如变形虫的状态。形态上的改变也伴随有IBA-1 和CD11b 表达的增加。
图1:小胶质细胞活化的代表示意图(改编自 Pinto et al., 2020)。在响应病原体或脑中内环境稳态改变时,小胶质细胞活化会引起形态和分子的改变。活化的细胞会部分地收缩其过程,从网状到中间的‘浓密’,到最终的具有吞噬功能、状如变形虫的状态。形态上的改变也伴随有IBA-1 和CD11b 表达的增加。
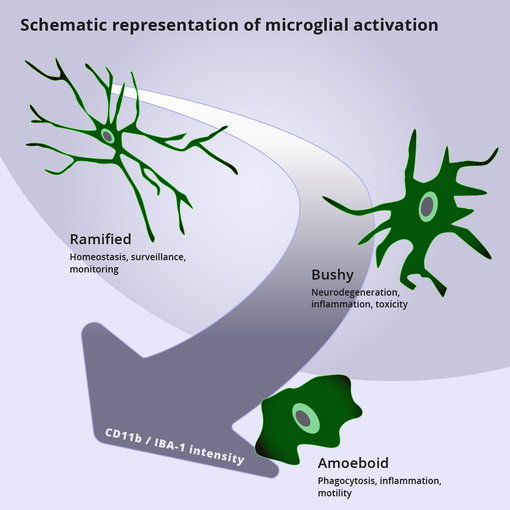
在响应传染性病原体、有害的蛋白质聚集体(例如Aβ,α-突触核蛋白,亨廷顿蛋白突变体,朊病毒)或肿瘤细胞时,小胶质细胞可以发起神经炎症反应(Hickman et al., 2018)。强烈的形态和分子的改变也伴随着小胶质细胞的活化(图1)(Pinto et al., 2020)。受刺激后,小胶质细胞会收缩并强化其过程,从网状到中间的‘浓密’状,到最终转换为变形虫样的状态(Yang et al., 2016; Pinto et al. 2020) (图2)。小胶质细胞的活化引起了IBA-1、MHCII、CD68 和CD11b,也称为整合素α M(ITGAM)表达的增加(Pinto et al., 2020, Jurga et al., 2020)。
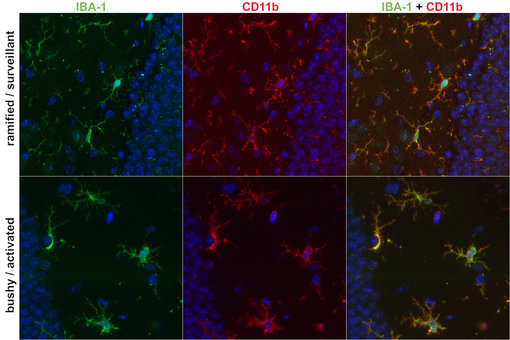
图2:通过IBA-1和CD11b染色识别小胶质细胞中形态的变化。野生型小鼠脑中的网状小胶质细胞(上行)。受病毒感染的FFPE小鼠脑中具有‘浓密’状的小胶质细胞(下行)。用大鼠抗-CD11b (cat. no. HS-384 117, 稀释比例1:100, 红色) 和豚鼠抗-IBA-1 (cat. no. HS-234 004, 1:500, 绿色)抗体,对经福尔马林固定、石蜡包埋(FFPE)的小鼠脑切片进行免疫组化染色。细胞核由DAPI 染色可见为蓝色。
图2:通过IBA-1和CD11b染色识别小胶质细胞中形态的变化。野生型小鼠脑中的网状小胶质细胞(上行)。受病毒感染的FFPE小鼠脑中具有‘浓密’状的小胶质细胞(下行)。用大鼠抗-CD11b (cat. no. HS-384 117, 稀释比例1:100, 红色) 和豚鼠抗-IBA-1 (cat. no. HS-234 004, 1:500, 绿色)抗体,对经福尔马林固定、石蜡包埋(FFPE)的小鼠脑切片进行免疫组化染色。细胞核由DAPI 染色可见为蓝色。
在响应由一氧化氮合成酶(iNOS)诱导合成的一氧化氮(NO)时,会引起小胶质细胞中CD11b的表达(Roy et al., 2006)。CD11b同CD18一起,又名为整合素β链-2,可形成整合素补体受体3(CR3),此受体会参与粘附过程、病原体的吞噬消除反应、炎症和耐受性响应的诱导、以及平行和下游宿主防御通路的调控(Jurga et al., 2020; Lamers et al., 2021)。CR3的表达并不仅限于小胶质细胞,也被发现可在中性粒细胞或者其它骨髓细胞包括巨噬细胞、单核细胞和嗜酸性粒细胞中表达(Lamers et al., 2021)。
K18-hACE2小鼠经鼻内感染Sars-CoV-2可引起致命疾病并伴有中枢神经系统严重炎症(Kumari et al., 2021)。同SARS-CoV-1 相似(Netland et al., 2008),SARS-CoV-2在脑中传播很快,在受感染的K18-hACE2小鼠嗅球和脑中可检测到高病毒滴度(Kumari et al. 2021)。病毒感染引起小胶质细胞活化,也伴随有CD11b和IBA-1表达的增加(图3)。此外,CD11b染色揭示了CD11b高中性粒细胞或者单核细胞浸润。
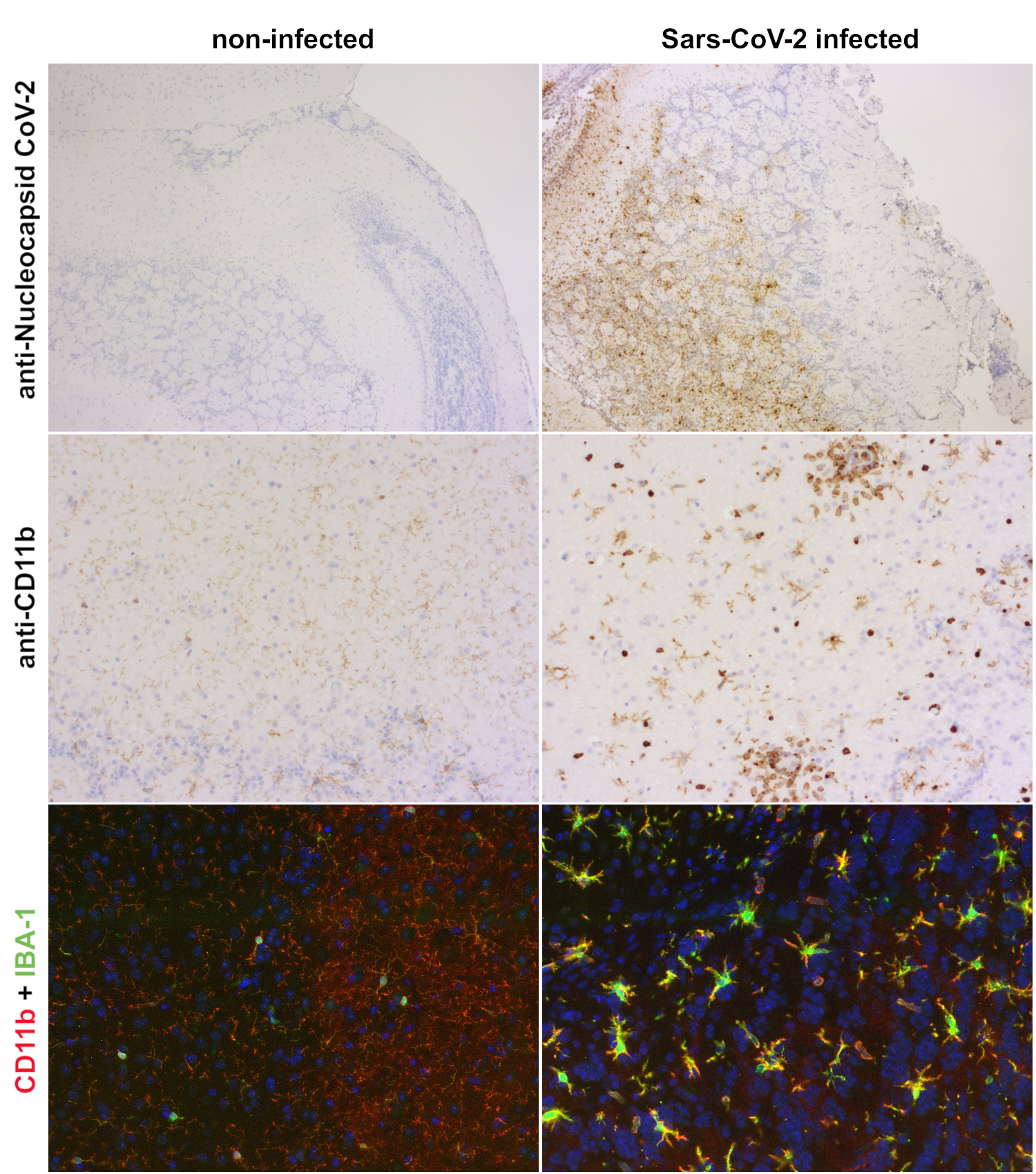
图3:感染Sars-CoV-2的hACE2 转基因小鼠脑中小胶质细胞的活化。第1行:使用抗-核衣壳蛋白CoV-2 (HS-452 011, 1:1000, DAB),在K18 hACE2 转基因小鼠的嗅球中检测到了感染了Sars-CoV-2 病毒的细胞(右列)。而在未感染的对照K18 hACE2转基因小鼠中没有检测到病毒蛋白(左列)。第2行:CD11b抗体染色揭示了在感染Sars-CoV-2的小鼠脑中,CD11b 在小胶质细胞中的表达以及CD11b阳性白血球浸润增加,而未感染的对照小鼠小胶质细胞中只检测到了少量CD11b (HS-384 117, 1:200, DAB)。第3行:使用CD11b (HS-384 117, 稀释比例1:100, 红色) 和IBA1 (HS-234 004, 1:500, 绿色) 进行免疫组化染色,可在感染Sars-CoV-2的小鼠嗅球小胶质细胞中识别CD11b和IBA-1的上调表达。小胶质细胞活化也引起了形态的变化。
败血症仍然是重症监护室里一个很重要的临床挑战,它会导致多器官功能障碍,也会影响中枢神经系统(CNS)。而在败血症病人的白质中(Zrzavy et al., 2019)和重症脓毒症的小鼠模型中(Michels et al., 2020),也发现了促炎性小胶质细胞活化。败血症相关的脑病(SAE)和慢性痛也被认为是大脑炎症的后果(reviewed in Li et al., 2020)。为了模拟人体中系统性炎症,不同的动物模型已被开发,包括脂多糖(LPS)处理、盲肠结扎和穿刺(CLP)、还有腹膜污染和感染(PCI)(Seemann et al., 2017)。腹膜污染和感染(PCI)的小鼠模型脑中小胶质细胞活化可用CD11b来检测(图4)。
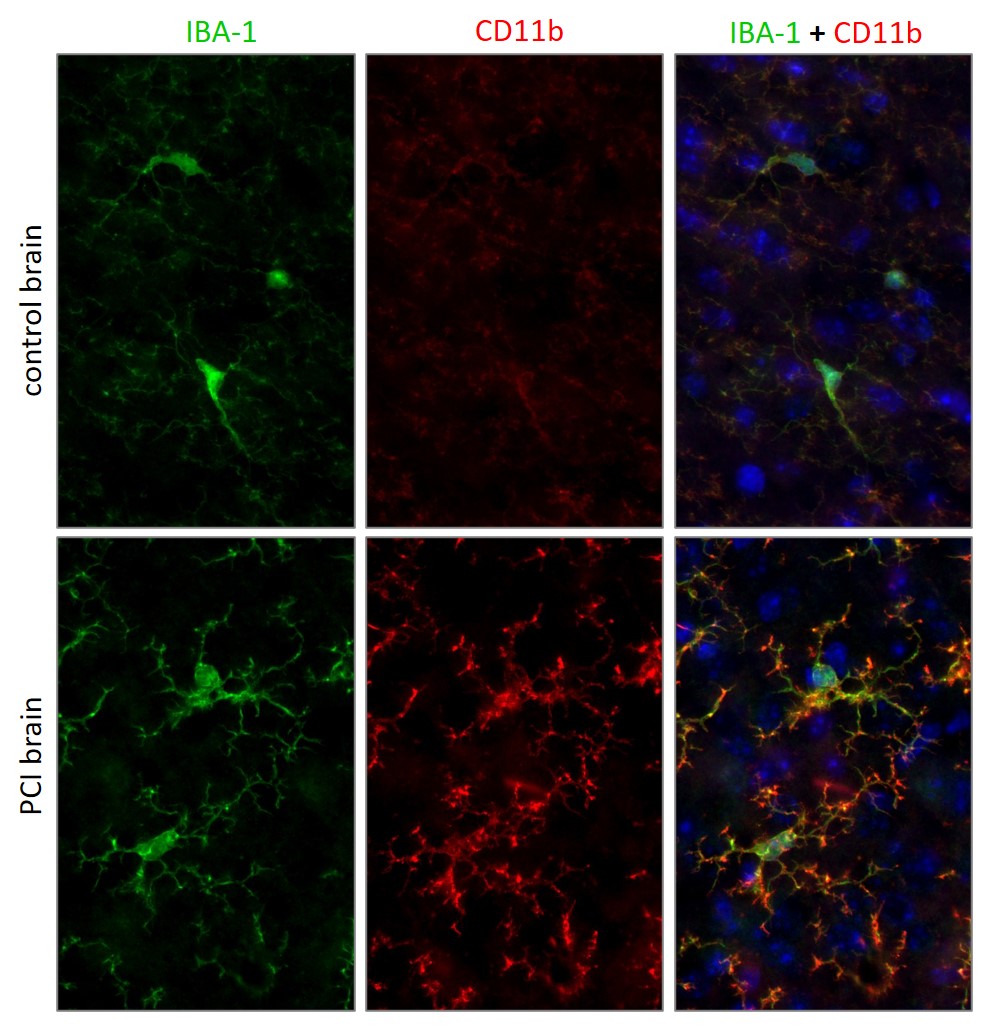
图4:来自野生型小鼠(对照脑;上行)和败血症小鼠模型(PCI脑;下行)的经PFA 固定的脑切片。使用大鼠抗-CD11b (cat. no. HS-384 117, 稀释比例1:500, 红色)和豚鼠抗-IBA1 (cat. no. HS-234 004, 稀释比例 1:500, 绿色)抗体,对切片进行免疫组化染色。细胞核由DAPI 染色可见为蓝色。
| Cat. No. | Product Description | Application | Quantity | Price | Cart |
|---|
| HS-384 008 | CD11b, rabbit, monoclonal, recombinant IgGrecombinant IgG mouse specific | WB IHC IHC-P IHC-Fr | 100 µl | US$420.00 | |
| HS-384 017 | CD11b, rat, monoclonal, purified IgG IgG human specific | WB IHC-P | 200 µl | US$420.00 | |
| HS-384 018 | CD11b, rabbit, monoclonal, recombinant IgGrecombinant IgG human specific | WB IHC-P IHC-Fr | 100 µl | US$420.00 | |
| HS-384 103 | CD11b, rabbit, polyclonal, affinity purifiedaffinity mouse specific | WB ICC IHC IHC-P | 200 µl | US$455.00 | |
| HS-384 117 | CD11b, rat, monoclonal, purified IgG IgG mouse specific | WB IHC IHC-P IHC-Fr | 200 µl | US$420.00 | |
| HS-384 117BT | CD11b, rat, monoclonal, purified IgG IgG, biotin mouse specific | WB IHC-P | 100 µg | US$470.00 | |
| HS-384 308 | CD11b, Guinea pig, monoclonal, recombinant IgGrecombinant IgG mouse specific | WB IHC IHC-P IHC-Fr | 100 µl | US$420.00 | |
| 375-0P | CD11c, control peptidecontrol peptide | 100 µg | US$110.00 | ||
| HS-375 003 | CD11c, rabbit, polyclonal, affinity purifiedaffinity mouse specific | WB IHC IHC-P | 200 µl | US$375.00 | |
| HS-375 004 | CD11c, Guinea pig, polyclonal, antiserumantiserum mouse specific | WB IHC IHC-P | 100 µl | US$355.00 | |
| HS-375 008 | CD11c, rabbit, monoclonal, recombinant IgGrecombinant IgG mouse specific | IHC IHC-P | 50 µg | US$420.00 | |
| HS-375 017 | CD11c, rat, monoclonal, purified IgG IgG mouse specific | IHC IHC-P IHC-Fr | 100 µg | US$420.00 | |
| HS-493 017 | CD39, rat, monoclonal, purified IgG IgG mouse specific | WB IHC IHC-P | 100 µg | US$420.00 | |
| HS-493 117 | CD39, rat, monoclonal, purified IgG IgG human specific | ICC IHC-P | 100 µg | US$420.00 | |
| HS-427 003 | CD45, rabbit, polyclonal, affinity purifiedaffinity human specific | WB ICC IHC-P | 50 µg | US$375.00 |
Certificates
ISO 9001 2015 Quality Management System and Green Lab Platinum certification level for sustaining laboratory processes.


Newsletter
Sign up for our newsletter and get the latest updates and news.