-
|
-
概览
• 组织驻留巨噬细胞的功能
• 形成组织驻留巨噬细胞身份的因素
• 鼠类组织驻留巨噬细胞的标记物
• 鼠类中枢神经系统中组织驻留巨噬细胞
• 小鼠肠道组织驻留巨噬细胞
• 小鼠肝脏组织驻留巨噬细胞
• 小鼠肺组织驻留巨噬细胞
• 小鼠脾脏组织驻留巨噬细胞
• 产品
• 参考文献
组织驻留巨噬细胞根据其位置和不同的基因表达谱而具有特殊的功能。它们参与组织发育过程(例如骨形成、脑及乳腺发育),组织内平衡(例如维持胰岛素敏感性、产热基因诱导、肝脂质平衡)和炎症的消除。与此相反,募集单核细胞会在组织中成为巨噬细胞,它被看作是应对炎症和病原体挑战的关键角色(Italiani et al., 2014)。
组织驻留巨噬细胞是免疫细胞的异质群体,以满足组织特异的和位置特异的功能(Davies et al., 2013)。组织驻留巨噬细胞的表现型由不同的因素塑造形成,它可被进一步分成起源、内在因素和外在因素(图1) (Blériot et al., 2020)。
起源。组织驻留巨噬细胞包含由三波不同前体产生的细胞,胚胎卵黄囊 (YS),胎肝和成人骨髓(BM) 来源的具有组织特异性比例的单核细胞 (Blériot et al., 2020)。YS来源的巨噬细胞(CD45+ CX3CR1bright F4/80bright)在血液中循环并在发育中的小鼠胚胎(E9.5-E10.5之间)中增殖,并从头部区域开始。只有从E12.5开始,胎肝造血开始活跃时,第二群体的F4/80low CD11bhigh细胞才会出现(Schulz et al., 2012)。
除小神经胶质细胞和一定比例的朗格汉斯细胞仍然保持卵黄囊起源以外,这些胎肝来源的巨噬细胞很大程度上取代了早期YS来源的巨噬细胞(Hoeffel et al., 2015)。
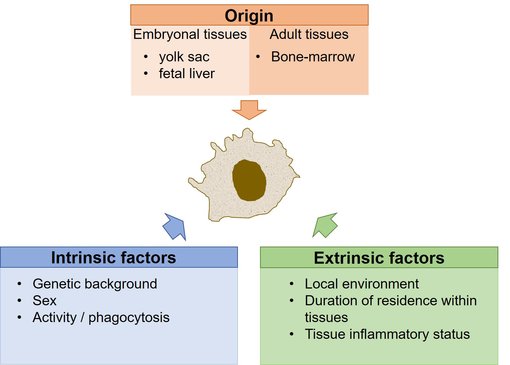
图1:形成组织驻留巨噬细胞身份的因素(改编自Blériot et al., 2020)。
除小神经胶质细胞和一定比例的朗格汉斯细胞仍然保持卵黄囊起源以外,这些胎肝来源的巨噬细胞很大程度上取代了早期YS来源的巨噬细胞(Hoeffel et al., 2015)。
图1:形成组织驻留巨噬细胞身份的因素(改编自Blériot et al., 2020)。
鼠类组织驻留巨噬细胞来源于胚胎前体,并在成年后通过自我更新保持其数量。三分之一的组织驻留巨噬细胞数量由成年BM来源的前体产生。因此,每个成年组织包含三个个体遗传上完全不同的巨噬细胞群。
内在因素。影响组织驻留巨噬细胞身份的内在因素包括遗传背景和性别。髓系基因突变与髓系白血病等疾病相关(Mueller et al., 2002)。性别身份会影响例如小神经胶质细胞的特性(Thion et al., 2018)。
外在因素。外在因素如何塑造组织驻留巨噬细胞身份的研究才刚刚开始。本地环境或者住所位置对巨噬细胞表现型具有主要的影响。由于组织不是完全相同的,他们包括不同细胞类型,器官特异性巨噬细胞也不是完全相同的。反而在同一组织中,位于独特亚组织位置的不同亚群的巨噬细胞中可能存在不同的程序(Blériot et al., 2020)。巨噬细胞在特定的局部环境中停留的时间会进一步影响巨噬细胞身份。新到来的单细胞来源的巨噬细胞与胚胎来源的巨噬细胞在相同器官中可共存。
鼠类组织驻留巨噬细胞来源于卵黄囊细胞,是典型的F4/80high巨噬细胞,例如皮肤中的朗格汉斯细胞、肝脏中的库普弗细胞和脑中的小神经胶质细胞(Schulz et al., 2012)。F4/80low CD11bhigh巨噬细胞是造血来源,并会不断地被BM衍生的祖细胞取代。BM衍生的祖细胞也可以取代传统的树突状细胞和在肾脏及肺中发现的一些F4/80high巨噬细胞(Schulz et al. 2012)。不论它们的起源,基因和细胞培养的研究表明,几乎所有巨噬细胞的主要谱系调节因子都是集落刺激因子-1受体,CSF1R(Wynn et al., 2013)。
表1:SYSY / HistoSure可提供的鼠类组织驻留巨噬细胞标记物。
| 标记物 | 抗体 | 应用 |
|
CD11b |
经常用于鉴别巨噬细胞和小神经胶质细胞,但是不能被所有组织驻留巨噬细胞表达。 | |
| CD11c | 可在树突状细胞、单核细胞、巨噬细胞、中性粒细胞和一小部分B细胞中表达。 | |
| CD45 | CD45 antibody, product no: HS-427 017, rat, mAb | 鼠类白血球的一种标记蛋白。CD45的表达水平可用来鉴别巨噬细胞亚群。 |
| CD163 | CD163 antibody, product no: HS-455 003, rabbit, pAb |
主要被组织驻留巨噬细胞表达。是中枢神经系统中血管周围巨噬细胞标记物。 |
| CD86 | CD86 antibody, product no: HS-466 003 | 传统的活化M1小神经胶质细胞和巨噬细胞的标记物。 |
| Chil3 | 在发生炎症时,分泌蛋白主要由巨噬细胞生成。 | |
| F4/80 |
F4/80 antibody, product no: HS-397 004, Guinea pig, serum |
鼠类巨噬细胞的标记物。在巨噬细胞亚群间表达有差异。 |
| IBA1 |
IBA1 antibody, product no: HS-234 004, Guinea pig, serum IBA1 antibody, product no: HS-234 013, rabbit, pAb IBA1 antibody, product no: HS-234 017, IBA1 rt mAb |
小神经胶质细胞和巨噬细胞的特定标记物。 |
| Tmem 119 |
Tmem 119 antibody, product no: 400 008, rabbit, mAb |
高度选择性小神经胶质细胞标记蛋白。 |
更多关于鼠类组织特异的巨噬细胞亚型如中枢神经系统、肠道、肝、肺及脾脏和用来鉴别这些亚型的标记物的详细信息,可在下面各部分找到。
小神经胶质细胞是中枢神经系统中关键的免疫效应细胞,分布遍及整个大脑。小神经胶质细胞作为病理性事件的感受器,对中枢神经系统的发育和维护非常重要。小神经胶质细胞来源于卵黄囊中红髓祖细胞,是CX3CR1high IBA-1pos F4/80pos CD11bpos CD45low (图 2) (Prinz et al., 2011)。Tmem 119是一种小神经胶质细胞高度特异的细胞表面标记物,不能被巨噬细胞或者其它免疫细胞或神经细胞类型表达(Bennett et al., 2016)。在炎症情况下,活体实验和体外实验表明CD11b在小神经胶质细胞中的表达会增加(Michels et al., 2020; Shen et al., 2017)。
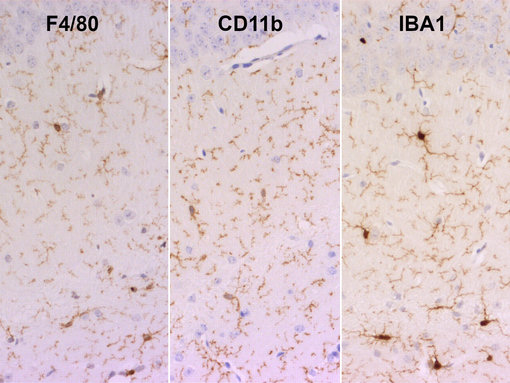
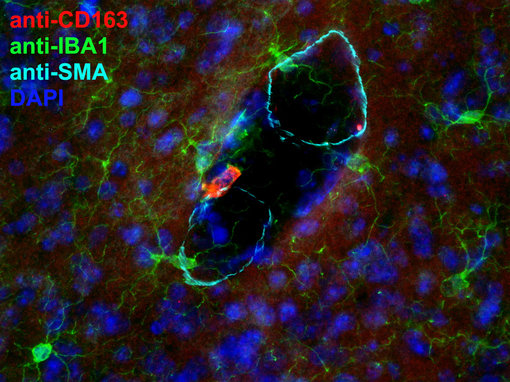
图2:使用兔抗-F4/80 (HS-397 008, 1:100)、大鼠抗-CD11b (HS-384 117, 1:100) 或者大鼠抗-IBA1 (HS-234 017, 10 µg/ml)抗体,对经福尔马林固定、石蜡包埋的小鼠脑切片中休眠的小神经胶质细胞进行染色。细胞核被苏木精复染为蓝色。
图3:使用兔CD163 (HS-455 003, 2 µg/ml; 红色),在经PFA固定的小鼠脑切片中,对血管周巨噬细胞进行间接免疫染色。CD163和IBA1 (Cat. 234 011, 绿色)可在血管周巨噬细胞而不是小神经胶质细胞中共定位。抗-α-平滑肌肌动蛋白(Cat. 449 004, 浅蓝)突出显示了血管边界。细胞核被DAPI复染为蓝色。
血管周巨噬细胞位于脑微血管的血管周围空间,是可长期存活的细胞,可忽略从外围循环的祖细胞得到的补充 (Kierdorf et al., 2019)。它们与本地免疫监视密切相关,是CX3CR1high CD163pos IBA-1pos F4/80pos CD11bpos CD45high (图 3) (Woong-Ki Kim et al., 2006; Prinz et al., 2011)
软脑膜和脉络膜丛巨噬细胞。其它位于中枢神经系统边界特定的巨噬细胞群体包括软脑膜和脉络膜丛巨噬细胞。同血管周巨噬细胞一起,它们被认为可支持和维护中枢神经系统相关结构的屏障功能,从而控制与中枢神经系统的代谢和抗原交换(Kierdorf et al., 2019)。
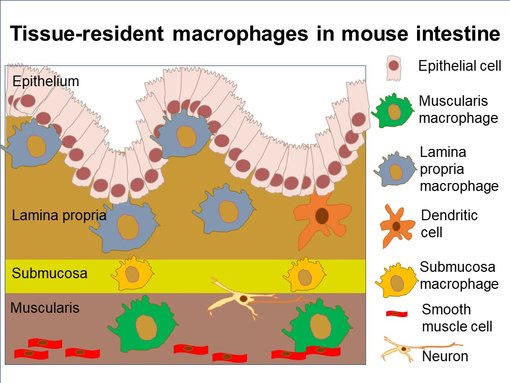
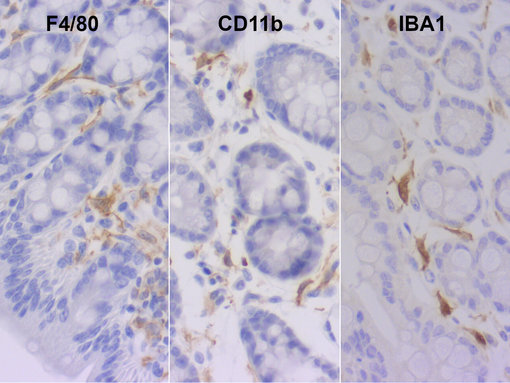
图4:小鼠肠道中亚组织位置(改编自 Viola et al., 2020a)。肠道巨噬细胞主要发现在固有层的绒毛里。黏膜下层的巨噬细胞与神经元和血管紧密相连。肌层巨噬细胞位于外肌层内。
图 5:在福尔马林固定、石蜡包埋的小鼠结肠中检测巨噬细胞和树突状细胞。使用大鼠抗-F4/80 (HS-397 017, 1:100) 对成熟巨噬细胞染色,大鼠抗-CD11b (HS-384 117, 1:100) 对树突状细胞和巨噬细胞染色,抗-大鼠IBA1 (HS-234 017, 10 µg/ml) 来检测活化的和休眠的巨噬细胞。细胞核被苏木精复染为蓝色。
肠道包含身体中最大的巨噬细胞池(Bain et al., 2018)。肠道巨噬细胞根据它们组织位置可被分为固有层巨噬细胞、黏膜下层巨噬细胞和肌层巨噬细胞(图4)。另一个巨噬细胞群可在黏膜相关的淋巴组织 (MALT)中发现,也就是派尔集合淋巴结和肠系膜淋巴结 (Viola et al., 2020a)。
肠道巨噬细胞在胚胎形成之前由卵黄囊或胎肝前体生成。胚胎出生后不久,大多数胚胎来源的巨噬细胞会快速由成年单核细胞来源的细胞替代 (Gross et al., 2015),主要位于固有层。然而,可长期存活且自我维持的巨噬细胞群仍在肠壁深处存在,与血管、黏膜下层和外肌层的肠和肌间神经元密切相关(Viola et al. 2020a)。长期存活的胚胎来源的巨噬细胞位置也很靠近派尔集合淋巴结和潘氏细胞(Ruder et al., 2020)。将来的Ly6Chigh单核细胞可通过MHCII 和CX3CR1的上调来区别于成熟的肠道巨噬细胞。完全成熟的组织驻留肠道巨噬细胞可表达F4/80、CD64、CD163 和CD206 (图 5) (Viola et al., 2020)。传统的树突状细胞 (cDC)也位于固有层,但缺乏巨噬细胞标记物F4/80、CD64和CX3XR1,从而可区别于巨噬细胞。小鼠和人体肠道固有层中的三种主要的传统树突状细胞亚型已被鉴别:类型1 CD103pos CD11bneg cDCs、类型2 CD103pos CD11bpos cDCs 和类型2 CD103neg CD11bpos cDCs (Sun et al., 2020)。
固有层巨噬细胞位于抵御细菌和食物抗原突破肠道屏障的第一防线,对维持病原体防御和口服耐受性之间的平衡起着重要的作用(Ruder et al., 2020)。固有层巨噬细胞具有很强的噬菌能力,并可表达高水平MHCII。固有层巨噬细胞也具有保持肠道上皮完整的功能,例如致病菌的清除和吞噬死亡细胞(Viola et al., 2020a)。
肌层巨噬细胞在转录水平和形态上是完全不同的,其特征是CD163、Retnla和Mrc1的上调(Viola et al., 2020)。此外,它们可表达M2相关基因如Arg1 和 Chil3(Ruder et al., 2020)。肌层巨噬细胞已被发现可调控肠道神经系统的神经元发育,并可直接与平滑肌细胞相互作用(Viola et al., 2020a)。
黏膜下层巨噬细胞专门支持肠道神经元和血管(Viola et al., 2020a)。黏膜下层神经元相关的巨噬细胞是可自我维持的,并可上调在巨噬细胞中更丰富的包括Fcrls、Mef2a、Hexb 和Gpr3等基因(Viola et al., 2020b)。
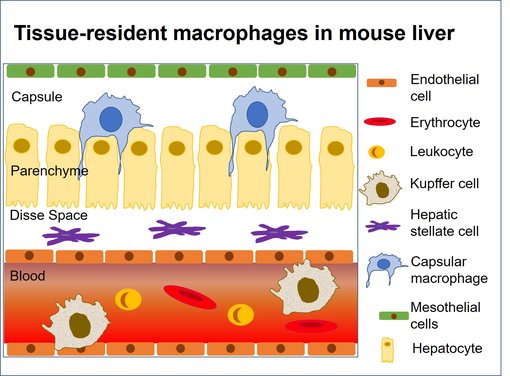
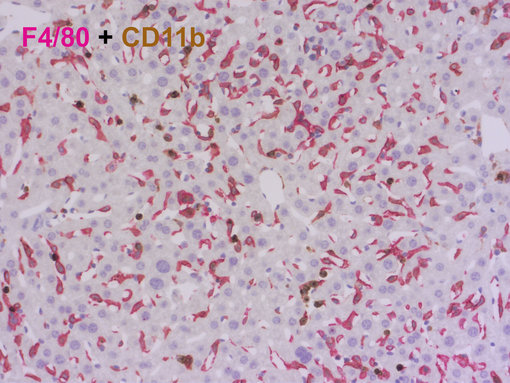
图 6:小鼠肝脏中的亚组织位置 (改编自 Blériot et al., 2020)。肝血窦中最丰富的巨噬细胞群体由库普弗细胞组成。
图 7:使用抗-F4/80 (HS-397 008, 1:100, AP-RED) 和抗-CD11b (HS-384 117, 1:100, DAB)进行发色体双重染色,可从 F4/80neg CD11bhigh 单核细胞和中性粒细胞中区分F4/80high CD11bneg 库普弗细胞。细胞核被苏木精复染为蓝色。
肝脏包含多种巨噬细胞群体:库普弗细胞,代表最丰富的肝脏巨噬细胞群体,而胶囊状的巨噬细胞,募集单核细胞来源的巨噬细胞和腹膜巨噬细胞(图 6) (Blériot et al., 2019)。
库普弗细胞大多数是F4/80pos CD11bneg Ly6cneg (Elchaninov et al., 2020),来源于胎肝单核细胞前体细胞,可自我更新,位于肝血窦,与血室直接接触(图 7)。库普弗细胞参与清除凋亡细胞和免疫复合物,识别和消除外来病原体,控制血液中铁、胆固醇和胆红素的平衡(Dou et al., 2020)。在小鼠肝脏中,CD163仅在大的库普弗细胞中有高水平表达(Elchaninov et al., 2019),然而,CD11b在骨髓来源的巨噬细胞中有更高水平的表达(Elchaninov et al., 2019)。
肝脏胶囊状巨噬细胞由成年循环单核细胞生成,是F4/80high CD11bpos CD11cpos Ly6Cpos (Blériot et al., 2019)。在炎症情况下,血液单核细胞来源的巨噬细胞 (F4/80neg to high CD11bpos to high CD11cneg Ly6Cpos to high)和成熟的腹膜巨噬细胞(F4/80high CD11bhigh CD11cneg Ly6Cpos)可以渗透进去肝脏(Blériot et al, 2019)。
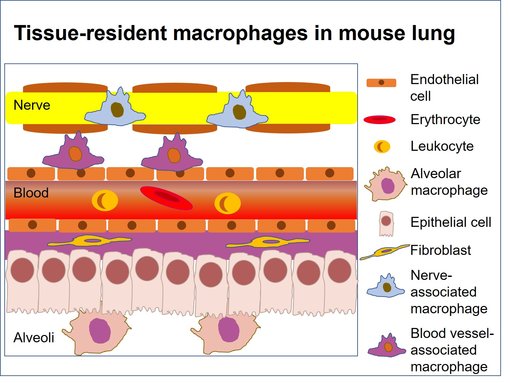
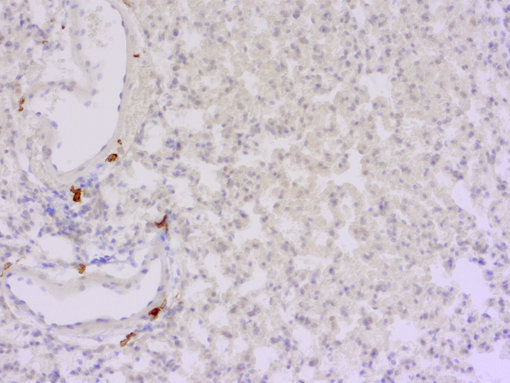
图8:小鼠肺中亚组织位置 (改编自Blériot et al., 2020)。小鼠肺包含两个主要的组织驻留巨噬细胞群体:肺泡中肺泡巨噬细胞和与血管或神经相关的间质巨噬细胞。
图 9: 对经福尔马林固定石蜡包埋的小鼠肺中兔 CD163 (HS-455 003, 1:250)进行染色发现,间质巨噬细胞中CD163-阳性,而肺泡巨噬细胞是CD163-阴性。细胞核被苏木精复染为蓝色。
肺髓室包括肺泡巨噬细胞、树突状细胞、组织单核细胞和间质巨噬细胞 (图 8) (Schyns et al., 2018)。
肺泡巨噬细胞来源于胚胎前体并具有自我更新的潜力(Akata et al., 2020)。肺泡巨噬细胞是专门负责循环利用表面活性剂分子,移除杂物,是CD11bneg SiglecF+pos CD11cpos CCR2neg CX3CR1neg (Schyns et al, 2018)。
间质巨噬细胞在胚胎出生之后来源于骨髓(Akata et al., 2020),并且可取代胚胎原始的间质巨噬细胞,它们在成年小鼠中优先定位于外围和血管周围区域(Tan et al., 2016)。间质巨噬细胞是CD11bpos SiglecFneg CD11clow CCR2low CX3CR1pos (Schyns et al., 2018) ,并且包括一些与血管相关的亚型(Chakarov et al., 2019) 或者导管相关的神经(Ural et al., 2020)。跟肺泡巨噬细胞相反,间质巨噬细胞可高度表达单核细胞相关基因,例如CD14、Csf1r 和CD163 (图 9) (Gibbings et al. 2017)。IBA1在间质巨噬细胞中含量高而在肺泡巨噬细胞中含量低(Donovan et al., 2018)。F4/80 可在胚胎原始巨噬细胞中表达,而在胚胎出生之后,其在骨髓来源的巨噬细胞中的表达是下调的(Tan et al., 2016)。
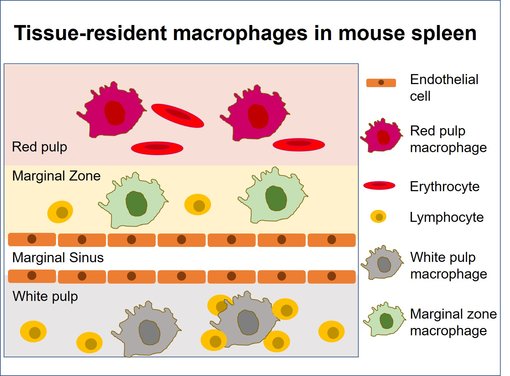
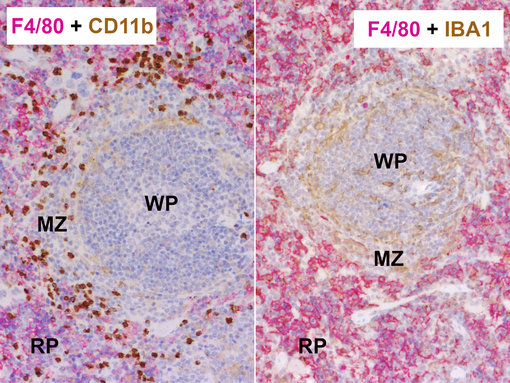
图10:小鼠脾脏中的亚组织位置(改编自 Blériot et al., 2020)。脾脏的组织驻留巨噬细胞是高度异质的,这是它们需要适应器官特定定位的结果:红髓巨噬细胞、白髓巨噬细胞和位于边缘地区的两种不同的巨噬细胞群体。
图11:巨噬细胞标记物兔抗-F4/80 (HS-397 008, 1:100, AP-RED, 红色) 和左图:大鼠抗-CD11b (HS-384 117, 1:100, DAB, 棕色) 或者右图:大鼠抗-IBA1 (HS-234 017, 10 µg/ml, DAB, 棕色) 的双重染色发现,经福尔马林固定石蜡包埋的小鼠脾脏中有不同的巨噬细胞亚群。细胞核被苏木精复染为蓝色。WP = 白髓;RP = 红髓;MZ = 边缘地区
鼠类脾脏包括四种巨噬细胞亚型:红髓巨噬细胞、白髓巨噬细胞和两种不同的边缘区巨噬细胞亚型(图10) (总结于 in Gonzales et al., 2018)
红髓巨噬细胞来源于胚胎YS-巨噬细胞和胚胎祖细胞,是F4/80pos VCAM1pos CD11blow CD163pos (图11)。红髓巨噬细胞参与衰老红血细胞的清理,在炎症刺激和寄生虫感染时履行免疫功能。
白髓巨噬细胞是F4/80neg CD68pos,会参与凋亡B细胞的吞噬(图11)。因此,它们可表达高水平的噬菌受体Mertk、Timd4和CD36。
边缘区巨噬细胞包括两种CD169pos SIGNR1pos 亚群, 又叫边缘区嗜金属巨噬细胞(MMMs)和边缘区巨噬细胞(MZMs)。MMMs 跟病毒的降解和清除有密切关系,扮演着抗原呈递细胞。MZMs 可高度表达清道夫受体MARCO,并和中枢耐受性有关。红髓巨噬细胞、MMMs和MZMs 染色显示IBA1阳性(Nakagawa et al., 2017)。而IBA1 染色在白髓巨噬细胞中却很少见(图11) (Donovan et al., 2018)。
| Cat. No. | Product Description | Application | Quantity | Price | Cart |
|---|
| HS-384 103 | CD11b, rabbit, polyclonal, affinity purifiedaffinity mouse specific | WB ICC IHC IHC-P | 200 µl | US$455.00 | |
| HS-384 117 | CD11b, rat, monoclonal, purified IgG IgG mouse specific | WB IHC IHC-P IHC-Fr | 200 µl | US$420.00 | |
| HS-375 003 | CD11c, rabbit, polyclonal, affinity purifiedaffinity mouse specific | WB IHC IHC-P | 200 µl | US$375.00 | |
| HS-375 004 | CD11c, Guinea pig, polyclonal, antiserumantiserum mouse specific | WB IHC IHC-P | 100 µl | US$355.00 | |
| HS-455 003 | CD163, rabbit, polyclonal, affinity purifiedaffinity mouse specific | WB IHC IHC-P | 200 µl | US$375.00 | |
| HS-427 017 | CD45, rat, monoclonal, purified IgG IgG mouse specific | IHC IHC-P IHC-Fr | 200 µl | US$420.00 | |
| HS-466 003 | CD86, rabbit, polyclonal, affinity purifiedaffinity mouse specific | WB IHC IHC-P IHC-Fr | 50 µg | US$375.00 | |
| HS-442 017 | Chil3, rat, monoclonal, purified IgG IgG mouse specific | WB ICC IHC IHC-P | 100 µg | US$420.00 | |
| HS-397 004 | F4/80, Guinea pig, polyclonal, antiserumantiserum | IHC IHC-P | 100 µl | US$355.00 | |
| HS-397 008 | F4/80, rabbit, monoclonal, recombinant IgGrecombinant IgG | WB IHC IHC-P | 100 µl | US$420.00 | |
| HS-397 017 | F4/80, rat, monoclonal, purified IgG IgG | WB IHC IHC-P IHC-Fr | 200 µl | US$420.00 | |
| HS-234 008 | IBA1, rabbit, monoclonal, recombinant IgGrecombinant IgG | WB ICC IHC IHC-P | 100 µl | US$425.00 | |
| HS-234 013 | IBA1, rabbit, polyclonal, affinity purifiedaffinity K.O. | WB IHC IHC-P | 200 µl | US$385.00 | |
| HS-234 017 | IBA1, rat, monoclonal, purified IgG IgG | WB ICC IHC IHC-P | 200 µl | US$420.00 | |
| 400 004 | TMEM119, Guinea pig, polyclonal, antiserumantiserum K.D. mouse specific | IHC IHC-P IHC-Fr | 100 µl | US$375.00 | |
| 400 008 | TMEM119, rabbit, monoclonal, recombinant IgGrecombinant IgG mouse specific | IHC IHC-P | 50 µg | US$420.00 |
Certificates
ISO 9001 2015 Quality Management System and Green Lab Platinum certification level for sustaining laboratory processes.


Newsletter
Sign up for our newsletter and get the latest updates and news.